Metabolismus
Durch Muskelkontraktionen werden Bewegungen realisiert. Hierbei wird für jede einzelne Muskelkontraktion Energie benötigt. Für jede Bewegung, so auch die repetitive Bewegung im Schwimmen, ist vor diesem Hintergrund die kontinuierliche Energiebereitstellung in Form von Adenosintriphosphat (ATP) von entscheidender Bedeutung. ATP steht der Muskelzelle jedoch nur in einem begrenzten Maße zur Verfügung (ca. 5 mmol ATP pro kg Muskelmasse) und muss entsprechend für eine andauernde muskuläre Beanspruchung fortlaufend wiederaufgebaut (resynthetisiert) werden. Dieser Prozess wird als Energiestoffwechsel bzw. Metabolismus bezeichnet. Der Energiestoffwechsel hat im Wesentlichen die Funktion ATP für den Vorgang der Muskelkontraktion zu generieren. Hierbei gibt es drei ATP-Resynthesewege, die in Abhängigkeit der Zeit und Belastungsintensität stehen (Hargreaves und Spriet, 2020).
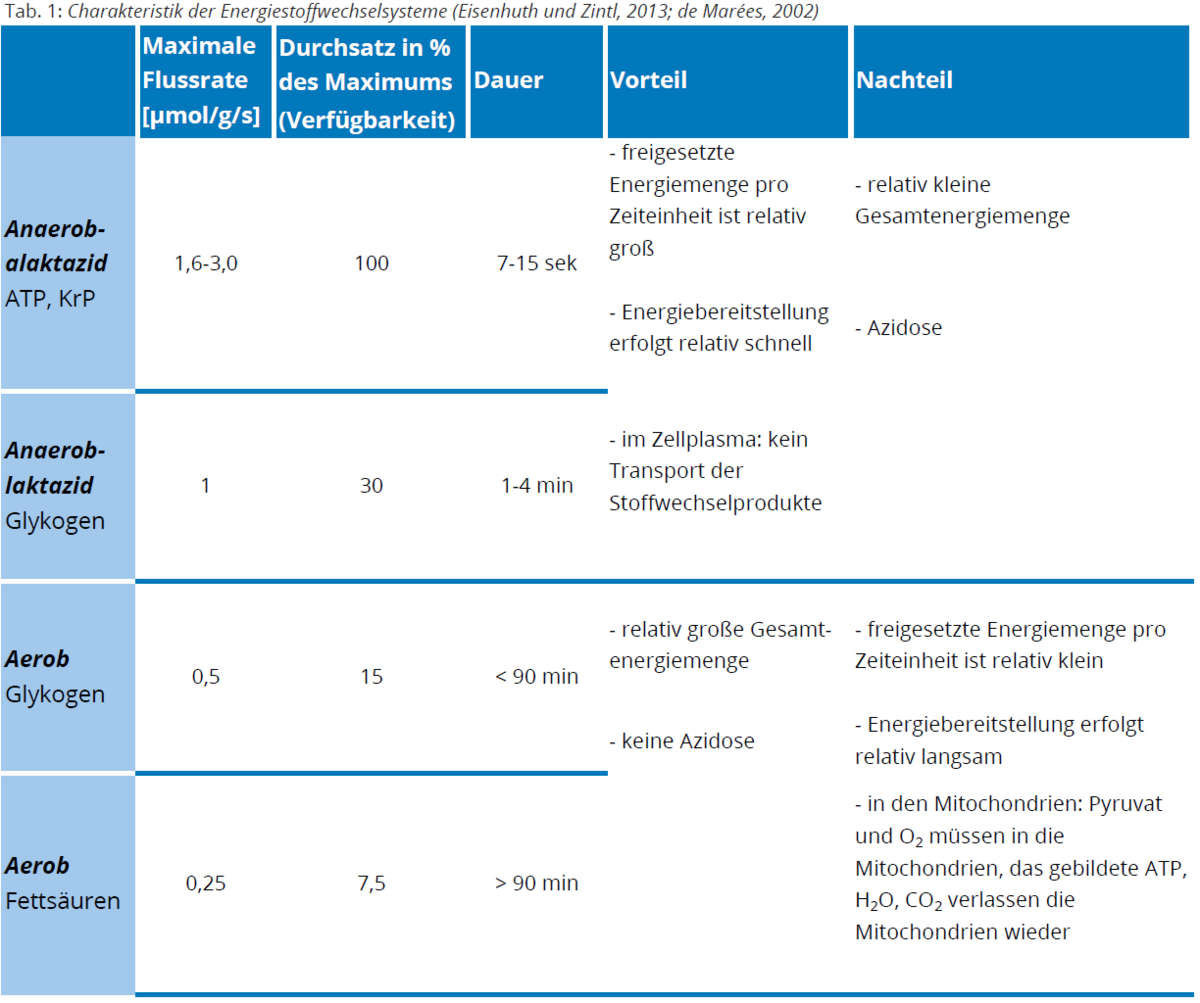
- Bei sehr kurzen Belastungszeiten mit einem hohen Energiebedarf erfolgt die Resynthese aus unmittelbar an der Muskelzelle lokalisierten und vergleichsweise kleinen Kreatinphosphatspeichern (Speicherkapazität zur ATP-Resynthese von bis zu 15 Sekunden, bei hohen Belastungsintensitäten; siehe 1 in Abb. 1).
- Mit einer geringeren ATP-Resyntheserate folgen die Energiebereitstellungswege der Glykolyse (Speicherkapazität zur ATP-Resynthese von 1 bis 4 Minuten, bei hohen bis mittleren Belastungsintensitäten; siehe 2 in Abb. 1) sowie
- der Oxidation (siehe 3 in Abb. 1) von Kohlenhydraten (Speicherkapazität zur ATP-Resynthese von ca. 90 Minuten, bei mittleren bis niedrigen Belastungsintensitäten) und Fetten (Speicherkapazität zur ATP-Resynthese von > 90 Minuten, bei niedrigen Belastungsintensitäten
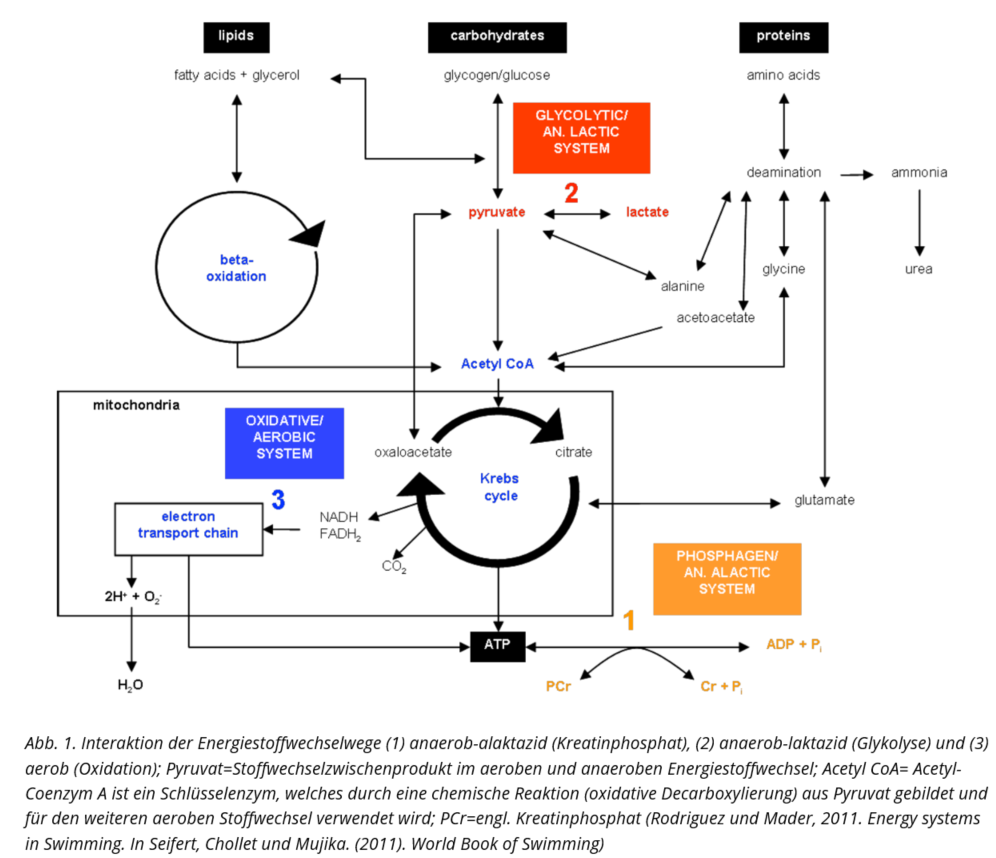
Generell laufen alle Energiebereitstellungswege parallel ab, es ändern sich nur deren Anteile am benötigten Gesamtenergiebedarf. Somit ist die Gesamtmenge der benötigten Energie gleich die Summe der einzelnen beschriebenen ATP-Resynthesewege (vgl. Abb. 2):
Etotal = Ealac + Elac + Eaer
Etotal=Gesamtenergiebedarf
Ealac=anaerob-alaktazide Energie (Kreatinphosphatspeicher)
Elac=anaerob-laktazider Energie (Glykolyse)
Eaer=aerobe Energie (Oxidation von Kohlenhydraten und Fetten)
Während die ersten beiden ATP-Resynthesewege (anaerob-alaktazid bzw. anaerob-laktazid) ohne den Verbrauch von Sauerstoff im Zellplasma (Zytoplasma) ablaufen, kann der letzte (bzw. die letzten beiden, wenn die aerobe Energiegewinnung aus Kohlenhydraten und Fetten unterschieden wird) nur unter der Verwendung von Sauerstoff (aerob) in den Mitochondrien erfolgen.
Eine besondere Stellung in diesem Gefüge nimmt der anaerob-laktazide Energiebereitstellungsweg, die Glykolyse, ein. Am Ende der Glykolyse entsteht nach einer Reihe enzymatischer Reaktionen Pyruvat, das nur in einem begrenzten Maße im Mitochondrium mit dem zur Verfügung stehenden Sauerstoff über den oxidativen Stoffwechselweg eliminiert werden kann. Bei einer Anhäufung von Pyruvat, die über der zu verstoffwechselnden Rate liegt (Akkumulation), wird Pyruvat über eine Reduktion (Aufnahme von zwei Wasserstoffionen) zu Laktat und Wasserstoffionen (H+) dissoziiert (Berg & Dickhuth, 2010). Die Akkumulation von Laktat und H+ wird vielfach mit einer Veränderung des Milieus, in dem eine Muskelkontraktion optimal ablaufen kann, beschrieben, wodurch nachfolgend die Leistungsfähigkeit der Muskulatur gehemmt wird. Als Ursache gilt die anfallende Laktatazidose, die jedoch als Ermüdungsursache kontrovers diskutiert wird (Böning & Maassen, 2008; Cairns, 2006; Wahl et al., 2009). Da der Laktatspiegel die Aktivität des anaeroben Stoffwechsels repräsentiert, wird die Laktatkonzentration hierzu repräsentativ, insbesondere in ausdauerdeterminierten Sportarten, verwendet.
Energiestoffwechselprofile im Schwimmen
Vor dem Hintergrund der bereits genannten Abhängigkeit der Energiestoffwechselaktivität zur Belastungsintensität und –zeit, ergeben sich für die Sportart Schwimmen Energiestoffwechselprofile, die das jeweilige Ausmaß der Beanspruchung der Energiestoffwechselwege wiedergeben. Nachfolgend sind diese für das Beckenschwimmen von 50m bis 1500m dargestellt.
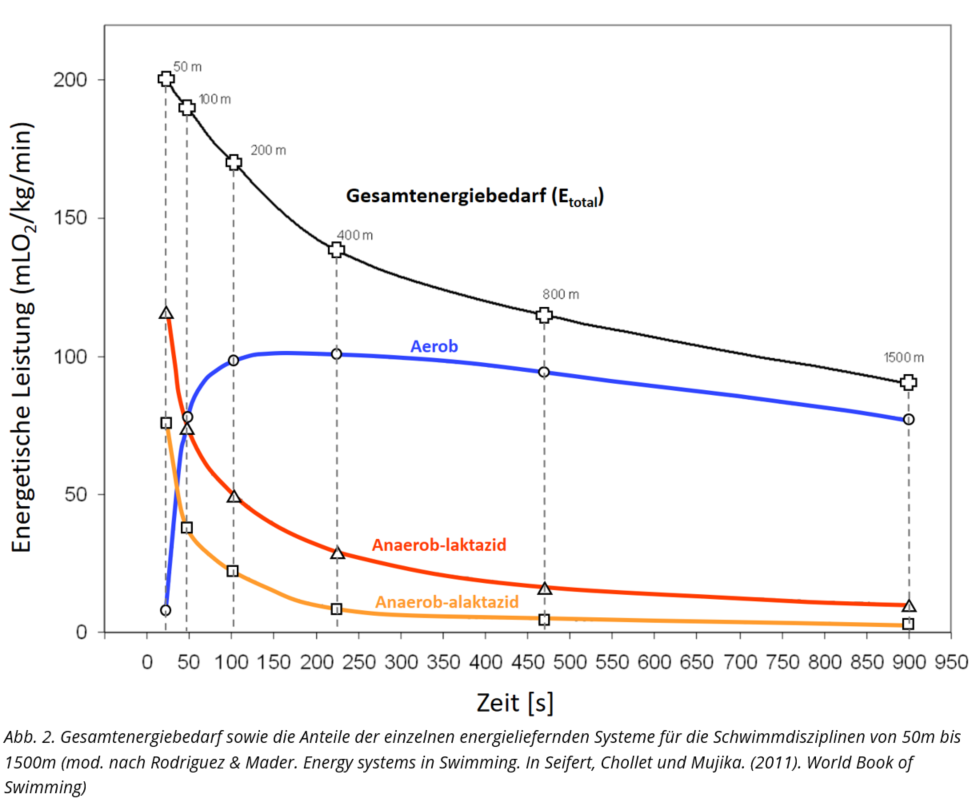
Für das Kraulschwimmen der Männer sind über die 50m- bis 1500m-Strecken mittels einer Simulationsrechnung die Energiestoffwechselprofile in der folgenden Übersicht dargestellt.
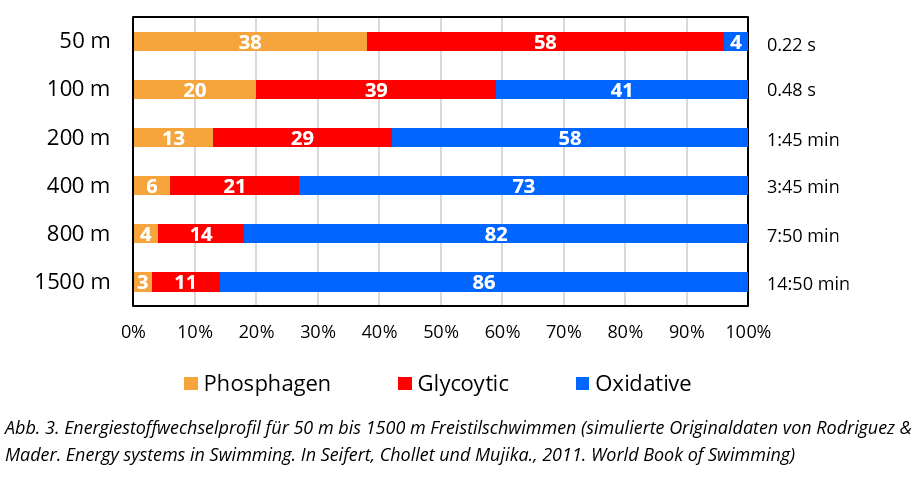
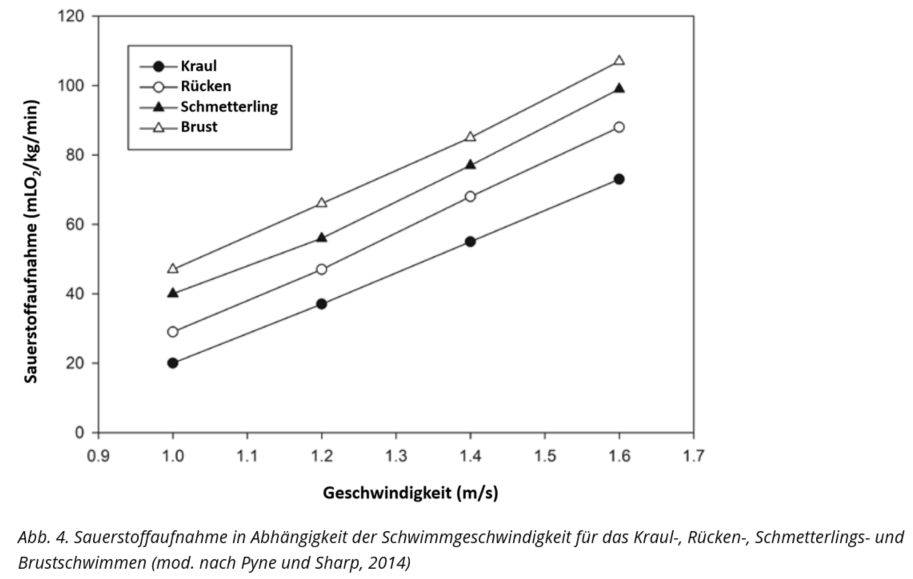
Eine vergleichbare Profilbildung wird für Frauen im Kraulschwimmen angenommen. Unter Berücksichtigung des Energiebedarfes, wiedergegeben anhand der Sauerstoffaufnahme in Abhängigkeit der Schwimmgeschwindigkeit (siehe Abb. 4), ist für die weiteren Schwimmlagen mit gewissen Abweichungen hinsichtlich des Energiestoffwechselprofils zu rechnen. Grundlegend ist das Kraulschwimmen als ökonomischster Schwimmstil zu benennen (siehe Abb. 4).
Einflussfaktoren der Laktatkonzentration
Für die Interpretation von Veränderungen der Laktatkonzentration unter Belastung ist das Verständnis vom Transport, der Verteilung sowie dem Verbrauch von Laktat nach dessen Produktion im Rahmen des anaeroben Energiestoffwechselweges entscheidend von Bedeutung. Früher wurde angenommen, dass Laktat aus der Muskelzelle nur entsprechend des Konzentrationsgefälles zwischen dem Intrazellular- und Extrazellularraum passiv diffundiert (Carré, 1993). Mittlerweile ist jedoch ein aktiver Transportmechanismus nachgewiesen worden, der akkumuliertes Laktat schneller aus der Muskelzelle befördert - die Monocarboxylat-Transporter (MCT). Über drei Wege kann auf der Basis dieses Transportsystems ein Laktattransport (engl.: lactate shuttle) erfolgen (Brooks, 2002; Brooks, 2009; Brooks & Hashimoto, 2007; Péronnet, 2010; Wahl et al., 2009):
1) intrazellulär: Das anfallende Laktat wird unmittelbar wieder in den Mitochondrien der Muskelzelle dem aeroben Energiestoffwechselweg zugeführt (intrazellulärer Laktat-Shuttle),
2) interzellulär: Das anfallende Laktat von glykolytischen Typ-I-Muskelfasern wird zu benachbarten oxidativen Typ-I-Muskelfasern transportiert und in den aeroben Energiestoffwechselweg eingespeist (Zell-Zell-Laktat-Shuttle),
3) extrazellulär: Das anfallende Laktat aus der Muskelzelle wird in das venöse Blut abgegeben, wo es nachfolgend u. a. in unbeanspruchter Muskulatur, dem Herzen oder der Leber wieder verstoffwechselt werden kann (extrazellulärer Laktat-Shuttle).
Damit bedingt neben der passiven Diffusion nur der dritte Laktat-Shuttle einen messbaren Anstieg der Laktatkonzentration im Blut. Die Konzentrationshöhe der MCTs geben dabei die Transportrate des Laktats zum einen aus der beanspruchten Muskelzelle und zum anderen in die Muskelzellen bzw. Organe, in denen das Laktat wieder verstoffwechselt wird, vor (Juel et al., 2003; Juel, 2004).
Dennoch ist es schwierig Mengenverhältnisse von MCTs mit Änderungen des Laktattransports und der Laktatkonzentration im Blut ins Verhältnis zu setzen, da die Laktatkinetik weiterhin von anderen Faktoren, wie der Ernährung, dem Trainingszustand oder der Hämodynamik beeinflusst wird (Juel, 2004; Wahl et al., 2009). In Erweiterung dessen wird der Verbrauch von Laktat als Metabolit von der Enzymdichte (z. B. Citrat-Synthase) in den Mitochondrien bestimmt. Vor diesem Hintergrund ist die Laktatkonzentration im Blut immer das Resultat aus der Wechselbeziehung zwischen der Produktionsrate, dem Transport, der Verteilung und dem Verbrauch. Dieser Sachverhalt wurde von Wahl et al. (2009) zusammenfassend wie folgt beschrieben:
„Blutlaktatkonzentrationen sind somit nicht das Resultat erhöhter Laktatbildung, sondern stellen das Ergebnis eines multifaktoriellen Prozesses dar. Trainingsbedingte Veränderungen der Blutlaktatkonzentration müssen daher als Ergebnis eines solchen multifaktoriellen Geschehens angesehen werden, bei dem der Laktattransport durch MCT eine wichtige Rolle spielt.“ (Wahl et al., 2009, S. 105).